| |
|
|
COMMON SPINAL CORD SYNDROMES |
|
Spinal cord disorders can extremely
important to recognize for two important reasons:
• Many
have treatable and
reversible causes if detected early
• If not detected and treated,
they can easily lead to permanent paralysis and disability
Relevant Spinal
Cord Anatomy
The well understood arrangement of tracts and cell columns in the spinal cord
frequently allows for precise neuroanatomic localization of signs and symptoms.
|
|
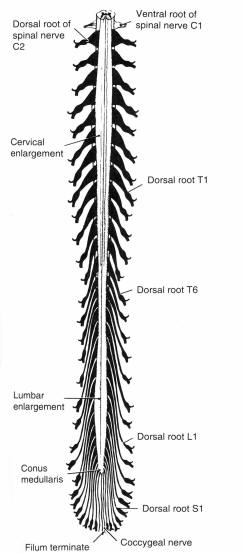 |
The spinal cord represents the caudal continuation of the lower brain stem
(medulla), beginning at the foramen magnum
and tapering over its 45-cm adult length to end in the
filum terminale, a narrow connective tissue band that anchors the
spinal cord to the coccyx. Over most of its course, the diameter of the spinal
cord is 1 cm or less, except for expansions in the cervical and lumbar spinal
cord which reflect the increased number of entering and exiting neurons relating
to the limbs. In cross section, the spinal cord is generally somewhat oval in
shape, wider in its transverse diameter, especially in its uppermost portions
and at the cervical and lumbar enlargements. Below the level of T12 the
substance of the cord tapers rapidly, forming the conically shaped
conus medullaris.
|
|
 |
|
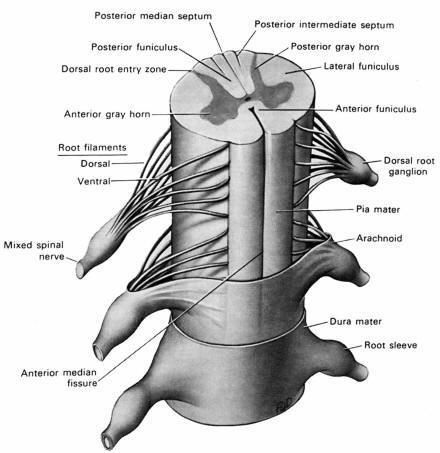
Examined in cross section, a central, distinctive butterfly-shaped
gray matter core is seen
surrounded by white matter tracts. A small,
ependyma-lined central canal runs the length of the cord and nearly all neurons
that cross from one side of the spinal cord to the other do so in the
anterior commissure that lies anterior to this
canal.
There is a general tendency in the spinal cord for
motor structures to be located anteriorly and sensory structures posteriorly.
Thus, the posterior gray matter of the spinal cord receives the dorsal (sensory)
roots, and the anterior gray of the spinal cord contains the anterior horn cells
(motor neuron cell bodies), which give rise to the anterior (motor) roots. The
descending motor (corticospinal) tracts are
located posterolaterally and the sensory white matter tracts are located both
anteriorly and posteriorly (dorsal and ventral
spinothalamic tracts and dorsal columns).
|
|
 |
|
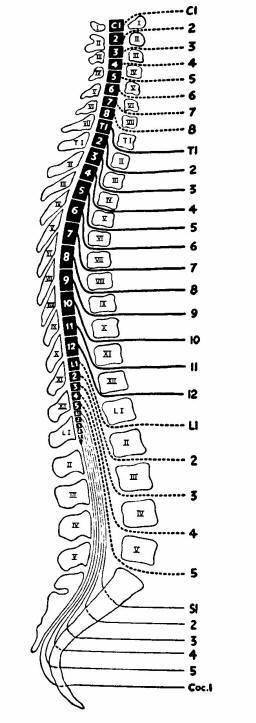
Each spinal nerve is named for its adjacent vertebral body. This leads to two
problems in nomenclature. Because there is an additional pair of spinal nerve
roots as compared to the number of vertebral bodies, the first seven spinal
nerves are named for the first seven cervical vertebrae, each nerve exiting
through the intervertebral foramen above its correspondingly named vertebral
body. The spinal nerve exiting below the level of C7, however, is referred to as
the C8 spinal nerve (the "extra" spinal root), though no eighth cervical
vertebra exists. Because of this "extra" nerve root, all subsequent roots exit
below the vertebral body for which they are named, beginning with T1.
The 8 cervical roots, 12 thoracic roots, 5 lumbar roots, 5
sacral roots, and 1 coccygeal root total 31 spinal nerve root pairs.
All of these contain both motor and sensory roots with the exception of C1,
which lacks a sensory component (explaining the absence of a "C1 dermatome").
After leaving the spine, the C5-T1 nerve roots from
the brachial plexus behind the clavicle. The 12 paired motor-sensory
thoracic roots continue as intercostal nerves. In the lower extremity, the
L1-S3 roots come together to form the lumbosacral plexus.
The other issue in spinal root nomenclature arises from the relative
positions of the spinal nerves with respect to their vertebral bodies. During
embryonic development, the spinal segments are closely aligned to their
corresponding vertebral segments. But later, the bony spinal column's downward
growth outpaces that of the spinal cord. This differential growth gives rise to
the appearance that the lower portion of the spinal cord has "ascended" in the
spinal canal relative to the vertebral column. Indeed, because
the adult spinal cord ends as the conus medullaris at
approximately the L1 level, the lumbar and sacral roots must plunge
downward below the termination of the spinal cord to find their respective
intervertebral foramina, forming the distinctive cauda
equina (horse tail). As a consequence, a pathologic process at the
level of the L4 vertebral body would be potentially in close proximity to both
the L4 nerve root and the other lower spinal roots of the cauda equina; however
a lesion at the L4 level cannot affect the spinal cord.
The vascular supply to the spinal cord consists of a single, larger
anterior spinal artery and two smaller
posterior spinal arteries. The anterior spinal
artery supplies the anterior two-thirds of the cord. The two smaller posterior
spinal arteries supply a wedge-shaped area constituting the posterior third of
the cord.
|
|
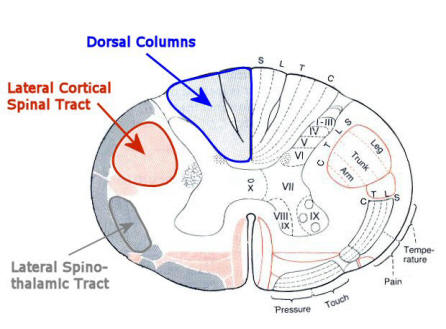
Functional Neuroanatomy of the Spinal Cord
While a large number of ascending and descending tracts have been identified
and mapped in the spinal cord, the three most important
of these in terms of neuroanatomic localization of spinal cord lesions are the
corticospinal tracts, spinothalamic tracts, and the dorsal columns
(see figure below).
|
|
|
|
The corticospinal tract arises from
neurons whose cell bodies are located in the precental gyrus, the primary motor
cortex). These neurons are known as the
upper motor
neurons. Once having crossed in the lower medulla, the axons of the
corticospinal tract move posteriorly to the posterolateral funiculi of the
spinal cord. Most of these neurons are destined to synapse on the anterior horn
cells located in the anterior gray of the spinal cord, the cell bodies of the
lower
motor neurons. These descending corticospinal tract fibers are
laminated in the spinal cord in a
clinically important arrangement, with fibers destined for the
lower limbs traveling more superficially in the cord and
fibers destined for upper limbs traveling more deeply in the cord.
Because they have already crossed, damage to these
corticospinal tract neurons in the spinal cord results in ipsilateral clinical
findings such as spastic weakness, increased deep tendon reflexes,
and a Babinski sign. When there is damage to the anterior horn cells (lower
motor neurons), ipsilateral clinical findings occur at the level of the affected
segments, including flaccid weakness, muscle wasting, decreased deep tendon
reflexes, and fasciculations.
There are two major ascending systems that transmit conscious sensory
information in the spinal cord: the spinothalamic
tracts and the dorsal columns
(a.k.a. posterior columns). The first order neurons of both of these afferent
systems begin as sensory structures situated in end organs (e.g., sensory
receptors in skin and stretch receptors in muscle). The cell bodies of the first
order neurons of these sensory pathways are located in the
dorsal root ganglia of the spinal nerves. These
ganglia are seen as distinctive prominences on the dorsal nerve roots just
proximal to the point where the anterior and dorsal branches join in the
intervertebral foramen to form the peripheral spinal nerve.
The spinothalamic tracts transmit pain and temperature sensation, commonly
tested at the bedside in the form of pinprick and cold sensation. As the axons
of these neurons enter the spinal cord, most rise one
or two levels before entering the dorsal gray of the spinal cord
where they synapse (explaining why a cord lesion at T3 may result in a level at
T5). They then cross immediately in the anterior commissure of the spinal cord and ascends in the anterolateral
funiculus as the lateral spinothalamic tract.
As a result, when the anterolaterally located spinothalamic tract is damaged in
the spinal cord, the patient experiences sensory
symptoms in the contralateral half of the body. This is contrary to
the case of injuries to the motor system described above, where the symptoms are
ipsilateral.
|
|
 |
|
Again, there is a clinically important lamination
(see figure above)of this tract where, like the corticospinal tract,
sensory neurons arising from the lower body travel more superficially in the
tract and neurons arising from higher levels travel more deeply in the tract.
This is important clinically as patients with external cervical spinal cord
disorders may first present with symptoms in their feet (as these fibers are
more superficial in the lateral spinothalamic tract than the fibers from the
upper extremities). |
The dorsal columns transmit vibratory and
proprioceptive information, commonly tested at the bedside by placing a
vibrating tuning fork on bone and by testing the patient's ability to detect
changes in joint position on passive motion. These neurons enter the spinal cord
via the dorsal root alongside pain and temperature neurons but, instead of
making an immediate synapse in the dorsal horn as do the latter type of neurons,
these axons enter the ipsilateral dorsal column
immediately, do not cross, and do not synapse until they reach the medulla.
Because this long, single neuron does not cross the midline until it passes
through the foramen magnum, a lesion involving one side
of the dorsal columns of the spinal cord causes ipsilateral loss of vibration
and joint position sense. Because the sensory modality of light touch
is transmitted through both the spinothalamic tracts and the dorsal columns,
light touch sensation is not completely lost unless both the spinothalamic and
dorsal column systems are affected. |
|
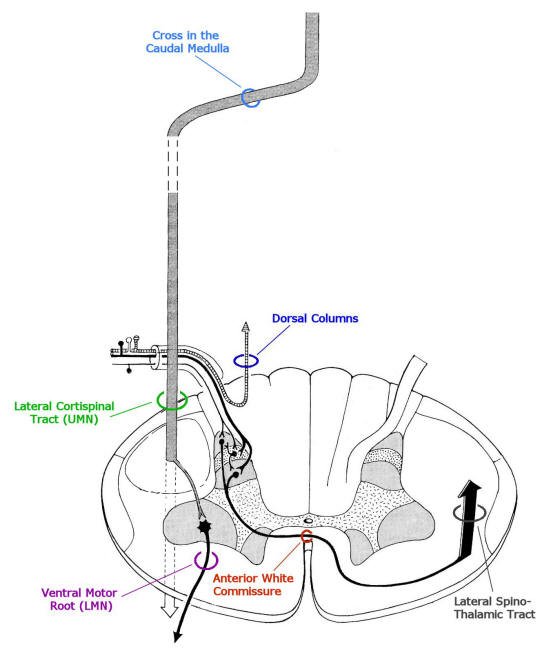
|
Above: Simplified view of the major sensory
and motor tracts used in localization of spinal cord disorders. Note on the
sensory side that the dorsal columns ascend ipsilaterally, while the lateral
spinothalamic tract crosses to the contralateral side and then ascends. On motor
side, both the lateral corticospinal tract (having crossed already above in the
lower medulla) and ventral motor root are ipsilateral.
|
|
Clinical Spinal Cord
Syndromes |
|
• Complete
Transection
• Hemisection
(Brown-Sequard)
• Combined Multisystem
Degeneration (Vitamin B12 deficiency)
• Anterior Horn Cell
Disease
• Central Cord
•
External Compression
• Anterior
Spinal Artery Occlusion |
|
|
|